




Abstract
The immune system has evolved to allow robust responses against pathogens while avoiding autoimmunity. This is notably enabled by stimulatory and inhibitory signals which contribute to the regulation of immune responses. In the presence of a pathogen, a specific and effective immune response must be induced and this leads to antigen-specific T-cell proliferation, cytokines production, and induction of T-cell differentiation toward an effector phenotype. After clearance or control of the pathogen, the effector immune response must be terminated in order to avoid tissue damage and chronic inflammation and this process involves coinhibitory molecules. When the immune system fails to eliminate or control the pathogen, continuous stimulation of T cells prevents the full contraction and leads to the functional exhaustion of effector T cells. Several evidences both in vitro and in vivo suggest that this anergic state can be reverted by blocking the interactions between coinhibitory molecules and their ligands. The potential to revert exhausted or inactivated T-cell responses following selective blocking of their function made these markers interesting targets for therapeutic interventions in patients with persistent viral infections or cancer.
1. Introduction
The immune system has evolved to allow robust responses against pathogens while avoiding autoimmunity. This is notably enabled by stimulatory and inhibitory signals which contribute to the regulation of immune responses. Positive costimulation is critical for the development of T-cell immune responses against foreign pathogens, while negative regulation is critical for the termination of immune responses, for peripheral tolerance, and to avoid inflammation-induced tissue damage [1–3].
When self/nonself antigens discrimination fails or when invading pathogens are not controlled, the immune system starts destroying cells and tissues of the body and consequently causes autoimmune diseases and chronic syndromes. In this regard, costimulatory and coinhibitory molecules are involved in regulating the initiation and termination of T-cell responses as well as spontaneous autoimmunity [3–5].
T-cell activation is determined by the presence of three distinct signals: (1) TCR-MHC class I and II interaction, (2) costimulatory molecules interaction, and (3) cytokines signaling. In the past, the dogma, based on initial observations, was that the integration of the distinct signals triggered T-cell activation, whereas the lack of complete positive signals led to tolerance or anergy [6–8]. More recently, the complexity of the model increased following the discovery of coinhibitory molecules triggering inhibitory signals. The functional outcome of costimulatory and coinhibitory molecules signaling is either enhancement or inhibition of TCR-mediated immune responses [9].
Over the past decade, four different families of costimulatory and coinhibitory molecules able to modulate TCR signaling have been identified: (1) B7-CD28 family including CD28, cytotoxic T-lymphocyte antigen-4 (CTLA-4; CD152), programmed death-1 (PD-1; CD279), inducible costimulatory molecule (ICOS; CD278), and B- and T-lymphocyte attenuator (BTLA; CD272) [1]; (2) CD2/signaling lymphocyte activation molecule (SLAM) family including SLAM (CD150), 2B4 (CD244), and CD48 [10, 11]; (3) Ig family including T-cell immunoglobulin mucin-3 (TIM-3) [12, 13], CD160 [14, 15], and lymphocyte-activation gene 3 (Lag-3) [16]; and (4) TNF-receptor superfamily including CD27 [17] (Figure 1).
Figure 1.
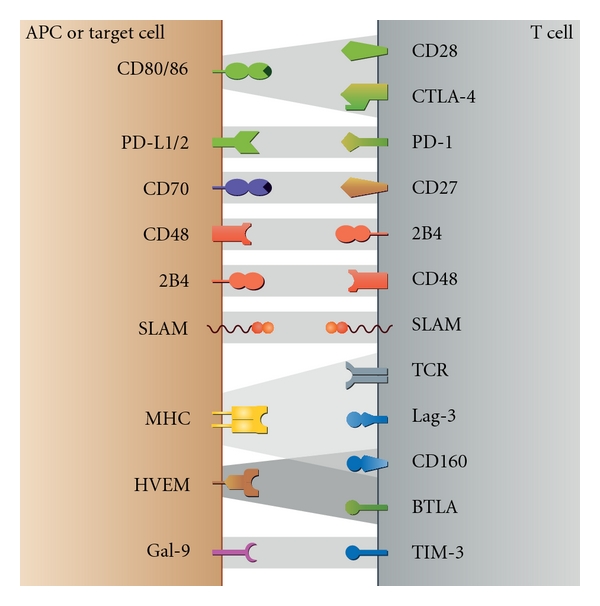
Regulatory molecules and their ligands. Schematic overview of the different costimulatory and coinhibitory molecules expressed by T cells (right panel) and association with their respective ligands expressed by antigen-presenting cells (APCs) or target cells (left panel). Coinhibitory molecules are color coded according to their relevant families. The four families of regulatory molecules include (1) B7-CD28 family including CD28, cytotoxic T-lymphocyte antigen-4 (CTLA-4; CD152), programmed death-1 (PD-1; CD279), inducible costimulatory molecule (ICOS; CD278), and B- and T-lymphocyte attenuator (BTLA; CD272); (2) CD2/signaling lymphocyte activation molecule (SLAM) family including SLAM (CD150), 2B4 (CD244), and CD48; (3) Ig family including T-cell immunoglobulin mucin-3 (TIM-3), CD160, and Lymphocyte-activation gene 3 (Lag-3); and (4) TNF-receptor superfamily including CD27.
In the presence of a pathogen, a specific and effective immune response must be induced and naïve T cells undergo activation upon encounter with their specific antigens [18, 19]. This leads to antigen-specific T-cell proliferation [20, 21], cytokines production, and induction of T-cell differentiation toward an effector phenotype [22] combined to survival signals [23, 24]. After clearance or control of the pathogen, the immune response must be terminated in order to avoid tissue damage and chronic inflammation [24, 25]. Two main mechanisms are involved in the contraction of the effector phase of immune responses, that is, either the inhibition of T-cell expansion [26] or the elimination of activated cells by apoptosis [27]. The latter is referred to as activation-induced cell death (AICD) [27, 28]. Direct inhibition of T-cell proliferation is induced via signals through coinhibitory molecules such as CTLA-4 or PD-1, while 2B4 and SLAM are considered to be critical in the regulation of AICD [29].
The role of coinhibitory molecules in regulating the immune system is also evidenced by severe autoimmune and lymphoproliferative diseases resulting from the lack or aberrant expression of these molecules [30].
2. Expression of Coinhibitory Molecules on Effector T Cells
T cells play an important role in the defense against infectious agents and tumors. Upon recognition of their cognate antigen, naïve T cells get activated and differentiate into effector cells [31]. This activation results in both phenotypic and functional changes that will determine the fate of effector T cells and the efficacy of the immune response [22]. Several studies have aimed to better define the profile of effector cells associated with the efficient control of infectious agents or tumors [32, 33]. While most studies focused on the differentiation state or functional profile of effector cells [34, 35], a lot of attention has been paid recently to the role of coinhibitory molecules [36].
SLAM family members, for instance, are immunomodulatory receptors associated with different functions including costimulation, cytokines production, and cytotoxic activity of immune cells (e.g., T cells or NK cells). Most members of this family serve as their own ligand on target cells or interact with molecules from the same family (e.g., CD48 and 2B4) [37, 38] and signal through a common messenger, that is, the SLAM-associated protein (SAP). SAP consists almost entirely of a single SH2 protein domain interacting with the cytoplasmic tail of SLAM and related receptors. One member of this family is SLAM and its signaling is involved in the induction and regulation of CD8 T-cell effector functions. In particular, SLAM is involved in the induction of IFN-γ production, cytotoxic activity, proliferation capacity, and activation-induced cell death (AICD) of activated CD4 and CD8 T cells [39–46]. In normal CD4 and CD8 T cells, SLAM enhanced TCR-mediated cytotoxicity and IFN-γ production [47], in contrast, T cells from SLAM-deficient mice showed increased IFN-γ production upon stimulation, thus supporting a negative role for SLAM in the regulation of IFN-γ production by effector T cells [10, 48, 49]. 2B4 (CD244), which is another important member of SLAM family, is a cell-surface glycoprotein structurally related to CD2-like molecules such as CD2, CD48, CD58, CD84, and Ly-9 [50] and seems to be particularly important for the cytotoxic effector function of CD8 T cells and NK cells [51–55].
Regarding the Ig family, CD160 which binds to classical and nonclassical HLA I molecules (i.e., HLA class Ia/b, HLA-C on NK cells) and Herpesvirus Entry Mediator (HVEM) [56], was identified not only on most NK cells and γδ T cells but also on a subset of CD8 αβ T cells [57, 58]. In NK cells, CD160 engagement induces cytotoxicity [59, 60] and has also been characterized as a marker of cytotoxic effector CD8 T cells [15, 61, 62]. However, the expression of CD160 on effector CD8 T cells is more controversial since no association between CD160 expression and perforin content was observed in CMV-specific CD8 T-cells [63] whereas, in HIV-1-infected patients, a CD160+CD8high effector T cell subset containing high amount of granzyme B (GrmB) has been described [64]. Furthermore, CD8 T cells expressing both 2B4 and CD160 were identified as a T-cell subset with a typical effector phenotype (i.e., CD27−CD45RA+CD56+CD57+) and expressing high levels of perforin and GrmB [62, 65].
Finally, TIM-3 is expressed at low levels on TH1 cells at a late stage of T-cell differentiation but not on TH2 cells, naïve T cells, B cells, macrophages, or dendritic cells. These evidences suggest that TIM-3 does not contribute to T-cell differentiation but has a role in the effector function of TH1 cells [66].
Also, during chronic viral infections, several inhibitory molecules are overexpressed on virus-specific T cells and this is associated with functional exhaustion. However, the expression of these molecules is also associated with the differentiation stage of T cells.
We have recently (Viganò et al., personal observation) performed a comprehensive investigation of the expression of PD-1, 2B4, CD160, CTLA-4, TIM-3, Lag-3, and SLAM on CD4 and CD8 T-cell subsets identified according to their differentiation state (i.e., Naïve, memory, effector/memory). These analyses showed that whereas virtually none of the coinhibitory molecules tested was present on naïve cells, those were present on memory T cells but at low levels, and more importantly, that effector/memory T cells expressed a significantly higher density coinhibitory molecules simultaneously (Figure 2).
Figure 2.

Expression of regulatory molecules following pathogen infection. Schematic overview of the pattern of expression of regulatory molecules. Following pathogen infection, key coinhibitory molecules are upregulated with different kinetics and play a role in regulating the development and the fate of effector T cells. In most cases, pathogens replication is controlled by the immune system leading to the contraction of effector T cells. Many different coinhibitory molecules, that is, PD-1, CTLA-4, BTLA, SLAM, and 2B4, play a role during this process. The remaining memory T cells (which are depending on the current models, derived either directly from naïve cells or from effector cells) express some coinhibitory molecules which depend on the type and biology of the pathogens. A hallmark of memory T cells is the lack of simultaneous expression of multiple coinhibitory molecules. Some regulatory molecules, however, are expressed by memory T cells, and this depends on the type of memory subset, that is, central or effector memory T cells. Conversely, when pathogens replication is not controlled, continuous stimulation of T cells, due to antigen persistence, prevents the full contraction of effector cells and leads to their functional exhaustion. A hallmark of exhausted effector cells is the simultaneous expression of several coinhibitory molecules. The simultaneous expression of these coinhibitory molecules is associated with their functional anergy.
3. Role of Coinhibitory Molecules in the Contraction of the Effector Phase of Immune Responses
The immune system is able to mount strong and efficient immune responses against pathogens without damaging organs [67]. This is notably achieved by the induction of the contraction and termination of the immune response after control or elimination of the infectious agent. During the contraction, the majority of effector T cells die, while remaining cells survive as memory cells [24, 25]. The elimination of effector cells mainly occurs via apoptosis and a number of pro- and antiapoptotic molecules were shown to be involved in this process [68]. The contraction of the immune response and the determination of T-cell fate depend on many transcription factors regulated during the course of the immune response [69]. These factors can be either induced or repressed by different signaling pathways provided, with different strength and kinetics, by costimulatory and coinhibitory molecules [70, 71].
One of the best-established mechanisms involved in the regulation of TCR signaling is the interaction between costimulatory (CD28) and coinhibitory (CTLA-4) molecules with CD80 or CD86 expressed by dendritic cells (DCs). Cross-linking of CD28 on T cells synergizes with TCR signaling to induce activation. Conversely, cross-linking of CTLA-4 induces an inhibitory signal which prevents T-cell activation [26, 72]. CTLA-4 is upregulated on activated T cells and, as a structural homologous to CD28 with higher affinity for CD80 or CD86, it competes with CD28 to inhibit TCR signaling [26].
PD-1 is another well-known regulatory molecule. It is expressed on activated CD4 and CD8 T-cells, NKT, B cells, and activated monocytes [1, 73], and its expression is induced by TCR- and BCR-mediated signaling [74]. The two PD-1 ligands (i.e., PD-L1 and PD-L2) differ in their expression pattern [75]. PD-L1 (B7-H1, CD274) is expressed by a broad array of cells (e.g., vascular endothelial cells, epithelial cells, muscles cells, hepatocytes) whereas PD-L2 (B7-DC, CD273) expression is restricted to hematopoietic cell types (i.e., DC, macrophages, mast cells) [76]. PD-1/PD-Ls pathway regulates the balance between stimulatory and inhibitory signals needed for effective immune responses against pathogens [77–80]. Engagement of PD-1 by PD-L1 leads to the inhibition of CD28-mediated costimulation and thus of TCR-mediated lymphocyte proliferation and cytokines secretion. The relative levels of expression of inhibitory (PD-Ls) and stimulatory (CD80/CD86) ligands by antigen-presenting cells (APC) can determine the extent of T-cell activation while PD-L1 expression on nonlymphoid tissues may determine the extent of effector immune responses at sites of inflammation [77]. Also member of the B7-CD28 family, BTLA, is an inhibitory receptor able to recruit phosphatases to dampen TCR signaling [81] through the interaction with HVEM expressed on naïve T and B cells. HVEM-BTLA signaling was shown to limit T-cell activity in vivo and to negatively regulate homeostatic expansion of CD4 and CD8 T cells [82]. Finally, HVEM can also interact with CD160, resulting in an inhibitory signaling dampening T-cell activation [83].
4. Functional Exhaustion and Loss of Effector Functions
During chronic viral infections such as HIV and HCV, several inhibitory molecules are overexpressed on virus-specific CD4 and CD8 T cells and this is associated with a state of functional deficiency also called functional exhaustion. Exhaustion is characterized by the progressive loss of T-cell functionality, leading ultimately to the deletion of exhausted T cells. The loss of the distinct T-cell functions occurs sequentially [84]. IL-2 production and T-cell proliferation potential are lost first. TNF-α production and cytotoxic capacity disappear later followed, ultimately, by the loss of IFN-γ production. Finally, deeply exhausted T cells are deleted via apoptosis [84].
The current hypothesis is that functional exhaustion occurs as a consequence of the attempt of the immune system to limit the magnitude of effector T-cell responses in order to safeguard against autoimmune responses and inflammatory damages. Nonetheless, this mechanism of protection may compromise effective immunity against persistent infectious agents and tumors [85].
Functional exhaustion occurs in the context of persistent high antigenic load and was first described in mice during chronic lymphocytic choriomeningitis virus (LCMV) infection [86] where LCMV-specific CD8 T cells persisted during the chronic phase of infection but lacked cytotoxic potential. Nonfunctional (i.e., anergic) antigen-specific CD8 T cells were also observed in the context of SIV [87], HIV [88], hepatitis B virus (HBV) [89], HCV [90, 91], and human T-lymphotropic virus 1 (HTLV1) [92] virus infection as well as in patients with persistent tumors [93]. However, mechanisms leading to exhaustion including the fundamental differences between exhausted cells and terminally differentiated cells or senescent (replication incompetent) cells remain unclear [94].
PD-1 was the first inhibitory receptor associated with immune exhaustion [95] in the seminal study performed in the LCMV model [96]. RNA microarray analyses of exhausted LCMV-specific CD8 T cells showed a marked upregulation of PD-1 expression [96]. Multiple studies have confirmed that high expression levels of PD-1 are associated with functional anergy and increased susceptibility to apoptosis [88, 97] in the context of human virus infections such as HIV [88, 91, 97–99], HCV [90, 91], HBV [89, 100], and also established tumors [101–105]. Of interest, it was reported that blockade of PD-1 signaling in vivo and in vitro resulted in the restoration of HIV-specific CD8 T-cell proliferation capacity and IL-2 production [96, 106]. However, the functional restoration by PD1/PD-L blockade was incomplete, and defects in CD8 T cells remained [96], suggesting the involvement of additional negative regulatory pathways in T-cell exhaustion [107, 108]. Analyses of global gene expression profiles of exhausted CD8 T cells identified the involvement of many coinhibitory receptors [109]. More recently, the severity of LCMV infection was associated to the number and the intensity of coinhibitory receptors expressed by virus-specific CD8 T cells [107].
Among these molecules, TIM-3, Lag-3, 2B4, CTLA-4, CD160, BTLA, KLRG1, CD305, and CD200R have been further investigated in the context of several human chronic virus infections and established tumors. In particular, the coexpression of TIM-3 and PD-1 was observed on both CD4 and CD8 T cells from patients with HIV [110] or HCV [111–113] chronic infections and correlated with T-cell exhaustion and diseases progression. In addition, TIM-3- and PD-1-expressing CD8 T cells represented a major population within tumor-infiltrating lymphocytes (TILs) in several murine models of cancer and in the blood of patients with advanced melanoma [114, 115]. In all cases, TIM-3/PD-1-expressing cells represented the most impaired population of CD8 T cells. Of note, the blockade of both molecules could restore CD8 T-cell effector functions (proliferation potential and cytotoxic capacity) of antigen-specific CD8 T cells and was associated with the control of tumor growth [110–113, 115].
CTLA-4 is another coinhibitory receptor upregulated in the context of chronic infections [91] and tumors [116]. It has been shown that CTLA-4 was overexpressed on CD4, but not CD8, T cells of SIV-infected macaques [117] and HIV-infected patients [118, 119]. Furthermore, the combination of CTLA-4 blockade and 4-1BB (CD137) activation enhanced tumor rejection by increasing T-cell infiltration, proliferation capacity, and cytokines production [120].
Of interest, BTLA was reported to be persistently expressed by melanoma-specific CD8 T cells, thus inhibiting their antitumor function [121]. On the other hand, BTLA expression on CD4 and CD8 T cells decreased during HIV infection and this was associated with CD4 T-cell differentiation and activation [122, 123]. Enhancing BTLA pathway may therefore represent an alternative therapeutic strategy to overcome immune activation during chronic HIV infection.
Lag-3 is an activation-induced cell-surface molecule, whose overexpression during chronic virus infection is also commonly associated with T-cell exhaustion and functional impairment. Blocking of Lag-3 alone failed in rescuing T-cell function or in decreasing plasma viremia during chronic LCMV infection [108], while blockade of both PD-1 and Lag-3 synergistically improved T-cell responses and decreased viral loads in vivo [107]. Elevated levels of Lag-3 and CTLA-4 were found in PD1+ CD4 T cells from HIV-infected patients [124] and in tumor-derived NY-ESO-1-specific CD8 T cells [125]. Functionality of these T-cell subsets was more impaired than in Lag-3−PD-1− or single Lag-3+ subsets [125].
SLAM family members are immunomodulatory receptors with a role in the regulation of costimulation, T-cell cytokines production, and cytotoxic activity. 2B4, which is a key molecule from this family, is involved in CD8 T-cell and NK-cell cytotoxicity. However, the proportion of 2B4+ CD8 T cells in HIV-infected patients correlated with immune activation of memory T cells and was increased in patients with progressive disease [126]. In addition, IFN-γ secretion and cytotoxic activity of 2B4+ CD8 T cells were significantly lower following stimulation with HIV as compared to influenza-derived antigens, respectively [127]. Furthermore, during infectious mononucleosis, the expression of SLAM and 2B4 on CD8 T cells correlated with severity of symptoms and viral loads [128].
The coexpression of molecules such as 2B4 and CD160, which have been related to potent cytolytic functions [62, 65], was associated with exhaustion and regulation of virus-specific CD8 and CD4 T cells in the context of chronic virus infections [83, 129]. A recent study showed a high frequency of CD8 T cells coexpressing PD-1, 2B4, CD160, KLRG1, LAG-3, and CTLA-4 in HCV infection. The coexpression of these molecules was associated with low levels of CD127 expression and correlated with impaired proliferation capacity [129].
The expression of another set of inhibitory molecules (i.e., PD-1, CTLA-4, CD305, and CD200R) has been investigated on CD4 T cells from HCV-infected patients. PD-1 and CTLA-4 were upregulated by HCV-specific CD4 T cells from patients with chronic infection, while CD305 and CD200R were upregulated in patients with cleared infection. Of note, the blockade of PD-Ls increased the expansion of CD4 T cells [130].
In the context of HIV infection, the presence of HIV-specific CD8 T cells coexpressing CD160, 2B4, and PD-1 but not Lag-3 was reported. The simultaneous expression of these molecules correlated with the level of virus replication and decreased cytokines production. The proliferative capacity was restored by blocking both PD-1/PD-L1 and 2B4/CD48 interactions [131]. Along the same line, another group showed that more than 30% of HIV-specific CD4 T cells expressed simultaneously PD-1, CTLA-4, and TIM-3, whereas less than 2% of CMV- or varicella-zoster virus-specific CD4 T cells coexpressed all three receptors. The coexpression of these molecules on HIV-specific CD4 T cells was more strongly correlated with the viral load compared with the expression of each receptor individually [132].
5. Potential Therapeutic Applications
The well-established immunosuppressive properties of coinhibitory molecules and the potential to revert exhausted or inactivated T-cell responses following selective blocking of their function made these markers interesting targets for therapeutic intervention in patients with persistent viral infections or cancer. To date, clinical and preclinical data are available for anti-CTLA-4 and anti-PD-1 blocking agents [87, 96, 133–136].
Initial human clinical trials assessing the effects of a blocking anti-CTLA-4 antibody demonstrated not only a reduction in tumor mass and clinical benefit in a minority of treated subjects but also an increase in systemic inflammation [137, 138]. Improvement in safety of these antibodies resulted in the recent approval by the U.S. Food and Drug Administration of a human monoclonal antibody against CTLA-4 (Ipilimumab, MDX-010, Yervoy) for the treatment of metastatic melanoma. In both early and late phase trials, Ipilimumab has demonstrated consistent activity against melanoma. However, serious (grade 3–5) immune-related adverse events occurred in 10–15% of patients. Thus, while providing a clear survival benefit, Ipilimumab administration requires careful patient monitoring combined to, sometimes, treatment with immune-suppressive therapy [133, 134]. In contrast, anti-CTLA-4 blockade failed to show benefit in terms of plasma viral load or survival in acutely or chronically SIV-infected macaques [139, 140]. Since CTLA-4 is preferentially upregulated on CD4 T cells and not on CD8 T cells [118], it might be possible that the blockade of anti-CTLA-4 induced an expansion and activation of CD4 T cells thus providing additional targets to HIV without significant improvement of CD8 T-cell functions.
Preclinical data showed how prevention of in vivo interactions between PD-1 and PD-L1 enhanced T-cell responses via the restoration of their ability to undergo proliferation, secrete cytokines, and lyse-infected cells and ultimately induce substantial reduction in viral loads. Of note, blockade of the PD-1/PD-L1 inhibitory pathway in vivo demonstrated a beneficial effect on CD8 T cells in mice that were lacking CD4 T-cell help [96]. This study identified a potentially effective immunotherapeutic strategy for chronic viral infections. This has then been further explored in nonhuman primates in a recent study evaluating the safety and immunomodulatory potential of an anti-PD-1 blocking antibody in SIV-infected macaques [87]. The treatment was well tolerated and led to a rapid increase in virus-specific CD8 T-cell responses with improved functional quality, both in peripheral and in GALT. PD-1 blockade also resulted in the expansion of virus-specific CD4 T cells, memory B cells, and higher titers of virus-specific antibodies. In contrast, one additional study showed an increase in CD4 T-cell activation and viral replication in mucosal sites [140]. Furthermore, a humanized anti-PD-1 monoclonal antibody (ONO-4538) is currently tested in a Phase 1 study in patients with recurrent or treatment-refractory cancer. Preliminary data support the safety, tolerability, and pharmacokinetic profile of a single-dose of the drug. In addition, preliminary evidences of antitumor activity were observed [135, 136].
There is currently a strong interest in the potential for clinical interventions targeting immunoregulatory networks to enhance immunity against cancer cells and persistent viruses or to boost the efficacy of preventive and therapeutic vaccines. The studies discussed previously have yielded promising results but have also highlighted important safety issues. This strongly indicates the importance to better understand mechanisms of immune regulation in order to exploit them for potential therapeutic applications.
6. Conclusion
Coinhibitory molecules are involved in maintaining the balance between the capacity to generate effector T cells able to control pathogens and the preservation of tolerance. During the development of immune responses, key coinhibitory molecules are upregulated with different kinetics and play a role in regulating the development and the fate of effector and memory T-cell responses. In most cases, pathogens replication is controlled by the immune system leading to the contraction of effector T cells. Many different coinhibitory molecules, that is, PD-1, CTLA-4, BTLA, SLAM, and 2B4, play a role during this phase. The remaining memory T cells express some coinhibitory molecules which depend on the type and biology of the pathogens and also on the level of differentiation. However, a hallmark of memory T cells is the lack of simultaneous expression of multiple coinhibitory molecules (Figure 2). Conversely, when pathogens replication is not controlled, continuous stimulation of T cells, due to antigen persistence, prevents the full contraction and leads to functional exhaustion of effector T cells. In contrast to memory T cells (see the aforementioned part), a hallmark of exhausted effector cells is the simultaneous expression of several coinhibitory molecules (Figure 2). The simultaneous expression of these coinhibitory molecules is associated with their functional anergy, also called exhaustion.
However, several evidences both in vitro and in vivo suggest that this anergic state can be reverted by blocking the interactions between coinhibitory molecules and their ligands. For this reason, coinhibitory molecules are now targets of preclinical and clinical studies aimed to identify new therapeutic strategies in the context of chronic infections and tumors. To date, only two coinhibitory molecules have been investigated in clinical trials, that is, PD-1 and CTLA-4, but recent evidences have underlined the importance of targeting multiple pathways in order to improve functional restoration. It is very likely that in the close future many additional targets will be assessed in preclinical and clinical studies.
In addition, while most studies focused their attention on the reversion of functional exhaustion, additional parallel strategies may be envisioned, such as the prevention of exhaustion in the context of therapeutic immunization.
Moreover, preventing/reverting exhaustion as a therapy for chronic conditions might be difficult to achieve notably for safety issues. On one hand, the prevention/reversion of exhaustion counteracts a physiological mechanism which is probably settled in order to avoid tissue damages and autoimmunity. On the other hand, restoration of functionality might not be sufficient since it will restore the functions of cells which failed to control the infection or to eliminate the pathogens. Therefore, it seems wise to plan to combine the functional restoration of T cells to other immunotherapeutic interventions.
Conflict of Interests
The authors declare that they have no conflict of interests.
Acknowledgment
This work was funded by the Swiss National Science Foundation.
References
- 1.Chen L. Co-inhibitory molecules of the B7-CD28 family in the control of T-cell immunity. Nature Reviews Immunology. 2004;4(5):336–347. doi: 10.1038/nri1349. [DOI] [PubMed] [Google Scholar]
- 2.Goodnow CC, Sprent J, Groth FdeS, Vinuesa CG. Cellular and genetic mechanisms of self tolerance and autoimmunity. Nature. 2005;435(7042):590–597. doi: 10.1038/nature03724. [DOI] [PubMed] [Google Scholar]
- 3.Bour-Jordan H, Esensten JH, Martinez-Llordella M, et al. Intrinsic and extrinsic control of peripheral T-cell tolerance by costimulatory molecules of the CD28/B7 family. Immunological Reviews. 2011;241(1):180–205. doi: 10.1111/j.1600-065X.2011.01011.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fife BT, Pauken KE. The role of the PD-1 pathway in autoimmunity and peripheral tolerance. Annals of the New York Academy of Sciences. 2011;1217(1):45–59. doi: 10.1111/j.1749-6632.2010.05919.x. [DOI] [PubMed] [Google Scholar]
- 5.Nurieva RI, Liu X, Dong C. Molecular mechanisms of T-cell tolerance. Immunological Reviews. 2011;241(1):133–144. doi: 10.1111/j.1600-065X.2011.01012.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Curtsinger JM, Lins DC, Mescher MF. Signal 3 determines tolerance versus full activation of naive CD8 T cells: dissociating proliferation and development of effector function. Journal of Experimental Medicine. 2003;197(9):1141–1151. doi: 10.1084/jem.20021910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mempel TR, Henrickson SE, Von Andrian UH. T-cell priming by dendritic cells in lymph nodes occurs in three distinct phases. Nature. 2004;427(6970):154–159. doi: 10.1038/nature02238. [DOI] [PubMed] [Google Scholar]
- 8.Curtsinger JM, Johnson CM, Mescher MF. CD8 T cell clonal expansion and development of effector function require prolonged exposure to antigen, costimulation, and signal 3 cytokine. Journal of Immunology. 2003;171(10):5165–5171. doi: 10.4049/jimmunol.171.10.5165. [DOI] [PubMed] [Google Scholar]
- 9.Leibson PJ. The regulation of lymphocyte activation by inhibitory receptors. Current Opinion in Immunology. 2004;16(3):328–336. doi: 10.1016/j.coi.2004.03.006. [DOI] [PubMed] [Google Scholar]
- 10.Wang N, Morra M, Wu C, et al. CD150 is a member of a family of genes that encode glycoproteins on the surface of hematopoietic cells. Immunogenetics. 2001;53(5):382–394. doi: 10.1007/s002510100337. [DOI] [PubMed] [Google Scholar]
- 11.Boles KS, Stepp SE, Bennett M, Kumar V, Mathew PA. 2B4 (CD244) and CS1: novel members of the CD2 subset of the immunoglobulin superfamily molecules expressed on natural killer cells and other leukocytes. Immunological Reviews. 2001;181:234–249. doi: 10.1034/j.1600-065x.2001.1810120.x. [DOI] [PubMed] [Google Scholar]
- 12.Zhu C, Anderson AC, Kuchroo VK. TIM-3 and its regulatory role in immune responses. Current topics in microbiology and immunology. 2011;350:1–15. doi: 10.1007/82_2010_84. [DOI] [PubMed] [Google Scholar]
- 13.Rodriguez-Manzanet R, Dekruyff R, Kuchroo VK, Umetsu DT. The costimulatory role of TIM molecules. Immunological Reviews. 2009;229(1):259–270. doi: 10.1111/j.1600-065X.2009.00772.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cai G, Freeman GJ. The CD160, BTLA, LIGHT/HVEM pathway: a bidirectional switch regulating T-cell activation. Immunological Reviews. 2009;229(1):244–258. doi: 10.1111/j.1600-065X.2009.00783.x. [DOI] [PubMed] [Google Scholar]
- 15.Anumanthan A, Bensussan A, Boumsell L, et al. Cloning of BY55, a novel Ig superfamily member expressed on NK cells, CTL, and intestinal intraepithelial lymphocytes. Journal of Immunology. 1998;161(6):2780–2790. [PubMed] [Google Scholar]
- 16.Huard B, Gaulard P, Faure F, Hercend T, Triebel F. Cellular expression and tissue distribution of the human LAG-3-encoded protein, an MHC class II ligand. Immunogenetics. 1994;39(3):213–217. doi: 10.1007/BF00241263. [DOI] [PubMed] [Google Scholar]
- 17.Denoeud J, Moser M. Role of CD27/CD70 pathway of activation in immunity and tolerance. Journal of Leukocyte Biology. 2011;89(2):195–203. doi: 10.1189/jlb.0610351. [DOI] [PubMed] [Google Scholar]
- 18.Goeken NE. Differential stimulatory requirements and regulation of naive and primed human lymphocytes. Human Immunology. 1984;10(4):251–263. doi: 10.1016/0198-8859(84)90090-9. [DOI] [PubMed] [Google Scholar]
- 19.Inaba K, Metlay JP, Crowley MT, Steinman RM. Dendritic cells pulsed with protein antigens in vitro can prime antigen-specific, MHC-restricted T cells in situ. Journal of Experimental Medicine. 1990;172(2):631–640. doi: 10.1084/jem.172.2.631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Henrickson SE, Mempel TR, Mazo IB, et al. In vivo imaging of T cell priming. Science Signaling. 2008;1(12):p. pt2. doi: 10.1126/stke.112pt2. [DOI] [PubMed] [Google Scholar]
- 21.Henrickson SE, Mempel TR, Mazo IB, et al. T cell sensing of antigen dose governs interactive behavior with dendritic cells and sets a threshold for T cell activation. Nature Immunology. 2008;9(3):282–291. doi: 10.1038/ni1559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kaech SM, Wherry EJ, Ahmed R. Effector and memory T-cell differentiation: implications for vaccine development. Nature Reviews Immunology. 2002;2(4):251–262. doi: 10.1038/nri778. [DOI] [PubMed] [Google Scholar]
- 23.Kaech SM, Wherry EJ. Heterogeneity and Cell-fate decisions in effector and memory CD8+ t cell differentiation during viral infection. Immunity. 2007;27(3):393–405. doi: 10.1016/j.immuni.2007.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mackay CR, Marston WL, Dudler L. Naive and memory T cells show distinct pathways of lymphocyte recirculation. Journal of Experimental Medicine. 1990;171(3):801–817. doi: 10.1084/jem.171.3.801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Badovinac VP, Harty JT. Programming, demarcating, and manipulating CD8+ T-cell memory. Immunological Reviews. 2006;211:67–80. doi: 10.1111/j.0105-2896.2006.00384.x. [DOI] [PubMed] [Google Scholar]
- 26.Chambers CA, Kuhns MS, Egen JG, Allison JP. CTLA-4-mediated inhibition in regulation of T cell responses: mechanisms and manipulation in tumor immunotherapy. Annual Review of Immunology. 2001;19:565–594. doi: 10.1146/annurev.immunol.19.1.565. [DOI] [PubMed] [Google Scholar]
- 27.Hildeman DA, Zhu Y, Mitchell TC, Kappler J, Marrack P. Molecular mechanisms of activated T cell death in vivo . Current Opinion in Immunology. 2002;14(3):354–359. doi: 10.1016/s0952-7915(02)00335-7. [DOI] [PubMed] [Google Scholar]
- 28.Arnold R, Brenner D, Becker M, Frey CR, Krammer PH. How T lymphocytes switch between life and death. European Journal of Immunology. 2006;36(7):1654–1658. doi: 10.1002/eji.200636197. [DOI] [PubMed] [Google Scholar]
- 29.Chen G, Tai AK, Lin M, Chang F, Terhorst C, Huber BT. Increased proliferation of CD8+ T cells in SAP-deficient mice is associated with impaired activation-induced cell death. European Journal of Immunology. 2007;37(3):663–674. doi: 10.1002/eji.200636417. [DOI] [PubMed] [Google Scholar]
- 30.Mueller DL. Mechanisms maintaining peripheral tolerance. Nature Immunology. 2010;11(1):21–27. doi: 10.1038/ni.1817. [DOI] [PubMed] [Google Scholar]
- 31.Butz EA, Bevan MJ. Massive expansion of antigen-specific CD8+ T cells during an acute virus infection. Immunity. 1998;8(2):167–175. doi: 10.1016/s1074-7613(00)80469-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Harari A, Enders FB, Cellerai C, Bart PA, Pantaleo G. Distinct profiles of cytotoxic granules in memory CD8 T cells correlate with function, differentiation stage, and antigen exposure. Journal of Virology. 2009;83(7):2862–2871. doi: 10.1128/JVI.02528-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Harari A, Dutoit V, Cellerai C, Bart PA, Du Pasquier RA, Pantaleo G. Functional signatures of protective antiviral T-cell immunity in human virus infections. Immunological Reviews. 2006;211:236–254. doi: 10.1111/j.0105-2896.2006.00395.x. [DOI] [PubMed] [Google Scholar]
- 34.Cellerai C, Perreau M, Rozot V, Enders FB, Pantaleo G, Harari A. Proliferation capacity and cytotoxic activity are mediated by functionally and phenotypically distinct virus-specific CD8 T cells defined by interleukin-7Rα (CD127) and perforin expression. Journal of Virology. 2010;84(8):3868–3878. doi: 10.1128/JVI.02565-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Harari A, Cellerai C, Enders FB, et al. Skewed association of polyfunctional antigen-specific CD8 T cell populations with HLA-B genotype. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(41):16233–16238. doi: 10.1073/pnas.0707570104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wherry EJ. T cell exhaustion. Nature Immunology. 2011;12(6):492–499. doi: 10.1038/ni.2035. [DOI] [PubMed] [Google Scholar]
- 37.Brown MH, Boles K, Van Der Merwe PA, Kumar V, Mathew PA, Barclay AN. 2B4, the natural killer and T cell immunoglobulin superfamily surface protein, is a ligand for CD48. Journal of Experimental Medicine. 1998;188(11):2083–2090. doi: 10.1084/jem.188.11.2083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Latchman Y, McKay PF, Reiser H. Cutting edge: identification of the 2B4 molecule as a counter-receptor for CD48. Journal of Immunology. 1998;161(11):5809–5812. [PubMed] [Google Scholar]
- 39.Cocks BG, Chan CCJ, Carballido JM, Yssel H, De Vries JE, Aversa G. A novel receptor involved in T-cell activation. Nature. 1995;376(6537):260–263. doi: 10.1038/376260a0. [DOI] [PubMed] [Google Scholar]
- 40.Howie D, Okamoto S, Rietdijk S, et al. The role of SAP in murine CD150 (SLAM)-mediated T-cell proliferation and interferon γ production. Blood. 2002;100(8):2899–2907. doi: 10.1182/blood-2002-02-0445. [DOI] [PubMed] [Google Scholar]
- 41.Howie D, Simarro M, Sayos J, Guirado M, Sancho J, Terhorst C. Molecular dissection of the signaling and costimulatory functions of CD150 (SLAM): CD150/SAP binding and CD150-mediated costimulation. Blood. 2002;99(3):957–965. doi: 10.1182/blood.v99.3.957. [DOI] [PubMed] [Google Scholar]
- 42.Henning G, Kraft MS, Derfuss T, et al. Signaling lymphocytic activation molecule (SLAM) regulates T cellular cytotoxicity. European Journal of Immunology. 2001;31(9):2741–2750. doi: 10.1002/1521-4141(200109)31:9<2741::aid-immu2741>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 43.Mehrle S, Schmidt J, Büchler MW, Watzl C, Märten A. Enhancement of anti-tumor activity in vitro and in vivo by CD150 and SAP. Molecular Immunology. 2008;45(3):796–804. doi: 10.1016/j.molimm.2007.06.361. [DOI] [PubMed] [Google Scholar]
- 44.Castro AG, Hauser TM, Cocks BG, et al. Molecular and functional characterization of mouse signaling lymphocytic activation molecule (SLAM): differential expression and responsiveness in Th1 and Th2 cells. Journal of Immunology. 1999;163(11):5860–5870. [PubMed] [Google Scholar]
- 45.Mikhalap SV, Shlapatska LM, Berdova AG, Law CL, Clark EA, Sidorenko SP. CDw150 associates with Src-homology 2-containing inositol phosphatase and modulates CD95-mediated apoptosis. Journal of Immunology. 1999;162(10):5719–5727. [PubMed] [Google Scholar]
- 46.Nagy N, Matskova L, Hellman U, Klein G, Klein E. The apoptosis modulating role of SAP (SLAM associated protein) contributes to the symptomatology of the X linked lymphoproliferative disease. Cell Cycle. 2009;8(19):3086–3090. doi: 10.4161/cc.8.19.9636. [DOI] [PubMed] [Google Scholar]
- 47.Henning G, Kraft MS, Derfuss T, et al. Signaling lymphocytic activation molecule (SLAM) regulates T cellular cytotoxicity. European Journal of Immunology. 2001;31(9):2741–2750. doi: 10.1002/1521-4141(200109)31:9<2741::aid-immu2741>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 48.Czar MJ, Kersh EN, Mijares LA, et al. Altered lymphocyte responses and cytokine production in mice deficient in the X-linked lymphoproliferative disease gene SH2D1A/DSHP/SAP. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(13):7449–7454. doi: 10.1073/pnas.131193098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cannons JL, Yu LJ, Hill B, et al. SAP regulates TH2 differentiation and PKC-θ-mediated activation of NF-κB1. Immunity. 2004;21(5):693–706. doi: 10.1016/j.immuni.2004.09.012. [DOI] [PubMed] [Google Scholar]
- 50.Valiante NM, Trinchieri G. Identification of a novel signal transduction surface molecule on human cytotoxic lymphocytes. Journal of Experimental Medicine. 1993;178(4):1397–1406. doi: 10.1084/jem.178.4.1397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Garni-Wagner BA, Purohit A, Mathew PA, Bennett M, Kumar V. A novel function-associated molecule related to non-MHC-restricted cytotoxicity mediated by activated natural killer cells and T cells. Journal of Immunology. 1993;151(1):60–70. [PubMed] [Google Scholar]
- 52.Nakajima H, Cella M, Langen H, Friedlein A, Colonna M. Activating interactions in human NK cell recognition: the role of 2B4-CD48. European Journal of Immunology. 1999;29(5):1676–1683. doi: 10.1002/(SICI)1521-4141(199905)29:05<1676::AID-IMMU1676>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 53.Schuhmachers G, Ariizumi K, Mathew PA, Bennett M, Kumar V, Takashima A. Activation of murine epidermal γδ T cells through surface 2B4. European Journal of Immunology. 1995;25(4):1117–1120. doi: 10.1002/eji.1830250440. [DOI] [PubMed] [Google Scholar]
- 54.Tangye SG, Cherwinski H, Lanier LL, Phillips JH. 2B4-mediated activation of human natural killer cells. Molecular Immunology. 2000;37(9):493–501. doi: 10.1016/s0161-5890(00)00076-6. [DOI] [PubMed] [Google Scholar]
- 55.Ostrowski SR, Ullum H, Pedersen BK, Gerstoft J, Katzenstein TL. 2B4 expression on natural killer cells increases in HIV-1 infected patients followed prospectively during highly active antiretroviral therapy. Clinical and Experimental Immunology. 2005;141(3):526–533. doi: 10.1111/j.1365-2249.2005.02869.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Agrawal S, Marquet J, Freeman GJ, et al. Cutting edge: MHC class I triggering by a novel cell surface ligand costimulates proliferation of activated human T cells. Journal of Immunology. 1999;162(3):1223–1226. [PubMed] [Google Scholar]
- 57.Maeda M, Carpenito C, Russell RC, et al. Murine CD160, Ig-like receptor on NK Cells and NKT cells, recognizes classical and nonclassical MHC class I and regulates NK cell activation. Journal of Immunology. 2005;175(7):4426–4432. doi: 10.4049/jimmunol.175.7.4426. [DOI] [PubMed] [Google Scholar]
- 58.Tsujimura K, Obata Y, Matsudaira Y, et al. Characterization of murine CD160+ CD8+ T lymphocytes. Immunology Letters. 2006;106(1):48–56. doi: 10.1016/j.imlet.2006.04.006. [DOI] [PubMed] [Google Scholar]
- 59.Buttner P, Mosig S, Lechtermann A, Funke H, Mooren FC. Exercise affects the gene expression profiles of human white blood cells. Journal of Applied Physiology. 2007;102(1):26–36. doi: 10.1152/japplphysiol.00066.2006. [DOI] [PubMed] [Google Scholar]
- 60.Le Bouteiller P, Tabiasco J, Polgar B, et al. CD160: a unique activating NK cell receptor. Immunology Letters. 2011;138(2):93–96. doi: 10.1016/j.imlet.2011.02.003. [DOI] [PubMed] [Google Scholar]
- 61.Maiza H, Leca G, Mansur IG, Schiavon V, Boumsell L, Bensussan A. A novel 80-kD cell surface structure identifies human circulating lymphocytes with natural killer activity. Journal of Experimental Medicine. 1993;178(3):1121–1126. doi: 10.1084/jem.178.3.1121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rey J, Giustiniani J, Mallet F, et al. The co-expression of 2B4 (CD244) and CD160 delineates an subpopulation of human CD8+ T cells with a potent CD160-mediated cytolytic effector function. European Journal of Immunology. 2006;36(9):2359–2366. doi: 10.1002/eji.200635935. [DOI] [PubMed] [Google Scholar]
- 63.Merino J, Ramírez N, Moreno C, Toledo E, Fernández M, Sánchez-Ibarrola A. BY55/CD160 cannot be considered a cytotoxic marker in cytomegalovirus- specific human CD8+ T cells. Clinical and Experimental Immunology. 2007;149(1):87–96. doi: 10.1111/j.1365-2249.2007.03387.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nikolova MH, Muhtarova MN, Taskov HB, et al. The CD160+ CD8high cytotoxic T cell subset correlates with response to HAART in HIV-1+ patients. Cellular Immunology. 2005;237(2):96–105. doi: 10.1016/j.cellimm.2005.01.012. [DOI] [PubMed] [Google Scholar]
- 65.Speiser DE, Colonna M, Ayyoub M, et al. The activatory receptor 2B4 is expressed in vivo by human CD8+ effector αβ T cells. Journal of Immunology. 2001;167(11):6165–6170. doi: 10.4049/jimmunol.167.11.6165. [DOI] [PubMed] [Google Scholar]
- 66.Monney L, Sabatos CA, Gaglia JL, et al. Th1-specific cell surface protein Tim-3 regulates macrophage activation and severity of an autoimmune disease. Nature. 2002;415(6871):536–541. doi: 10.1038/415536a. [DOI] [PubMed] [Google Scholar]
- 67.Hugues S, Fetler L, Bonifaz L, Helft J, Amblard F, Amigorena S. Distinct T cell dynamics in lymph nodes during the induction of tolerance and immunity. Nature Immunology. 2004;5(12):1235–1242. doi: 10.1038/ni1134. [DOI] [PubMed] [Google Scholar]
- 68.Taylor RC, Cullen SP, Martin SJ. Apoptosis: controlled demolition at the cellular level. Nature Reviews Molecular Cell Biology. 2008;9(3):231–241. doi: 10.1038/nrm2312. [DOI] [PubMed] [Google Scholar]
- 69.Hughes PD, Belz GT, Fortner KA, Budd RC, Strasser A, Bouillet P. Apoptosis regulators Fas and Bim cooperate in shutdown of chronic immune responses andprevention of autoimmunity. Immunity. 2008;28(2):197–205. doi: 10.1016/j.immuni.2007.12.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Iezzi G, Karjalainen K, Lanzavecchia A. The duration of antigenic stimulation determines the fate of naive and effector T cells. Immunity. 1998;8(1):89–95. doi: 10.1016/s1074-7613(00)80461-6. [DOI] [PubMed] [Google Scholar]
- 71.Salomon B, Bluestone JA. Complexities of CD28/B7: CTLA-4 costimulatory pathways in autoimmunity and transplantation. Annual Review of Immunology. 2001;19:225–252. doi: 10.1146/annurev.immunol.19.1.225. [DOI] [PubMed] [Google Scholar]
- 72.Collins AV, Brodie DW, Gilbert RJC, et al. The interaction properties of costimulatory molecules revisited. Immunity. 2002;17(2):201–210. doi: 10.1016/s1074-7613(02)00362-x. [DOI] [PubMed] [Google Scholar]
- 73.Greenwald RJ, Freeman GJ, Sharpe AH. The B7 family revisited. Annual Review of Immunology. 2005;23:515–548. doi: 10.1146/annurev.immunol.23.021704.115611. [DOI] [PubMed] [Google Scholar]
- 74.Nakae S, Suto H, Iikura M, et al. Mast cells enhance T cell activation: importance of mast cell costimulatory molecules and secreted TNF. Journal of Immunology. 2006;176(4):2238–2248. doi: 10.4049/jimmunol.176.4.2238. [DOI] [PubMed] [Google Scholar]
- 75.Ishida M, Iwai Y, Tanaka Y, et al. Differential expression of PD-L1 and PD-L2, ligands for an inhibitory receptor PD-1, in the cells of lymphohematopoietic tissues. Immunology Letters. 2002;84(1):57–62. doi: 10.1016/s0165-2478(02)00142-6. [DOI] [PubMed] [Google Scholar]
- 76.Latchman Y, Wood CR, Chernova T, et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nature Immunology. 2001;2(3):261–268. doi: 10.1038/85330. [DOI] [PubMed] [Google Scholar]
- 77.Freeman GJ, Long AJ, Iwai Y, et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. Journal of Experimental Medicine. 2000;192(7):1027–1034. doi: 10.1084/jem.192.7.1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Okazaki T, Honjo T. The PD-1-PD-L pathway in immunological tolerance. Trends in Immunology. 2006;27(4):195–201. doi: 10.1016/j.it.2006.02.001. [DOI] [PubMed] [Google Scholar]
- 79.Nishimura H, Nose M, Hiai H, Minato N, Honjo T. Development of lupus-like autoimmune diseases by disruption of the PD-1 gene encoding an ITIM motif-carrying immunoreceptor. Immunity. 1999;11(2):141–151. doi: 10.1016/s1074-7613(00)80089-8. [DOI] [PubMed] [Google Scholar]
- 80.Nishimura H, Okazaki T, Tanaka Y, et al. Autoimmune dilated cardiomyopathy in PD-1 receptor-deficient mice. Science. 2001;291(5502):319–322. doi: 10.1126/science.291.5502.319. [DOI] [PubMed] [Google Scholar]
- 81.Sedy JR, Gavrieli M, Potter KG, et al. B and T lymphocyte attenuator regulates T cell activation through interaction with herpesvirus entry mediator. Nature Immunology. 2005;6(1):90–98. doi: 10.1038/ni1144. [DOI] [PubMed] [Google Scholar]
- 82.Krieg C, Boyman O, Fu YX, Kaye J. B and T lymphocyte attenuator regulates CD8+ T cell-intrinsic homeostasis and memory cell generation. Nature Immunology. 2007;8(2):162–171. doi: 10.1038/ni1418. [DOI] [PubMed] [Google Scholar]
- 83.Cai G, Anumanthan A, Brown JA, Greenfield EA, Zhu B, Freeman GJ. CD160 inhibits activation of human CD4+ T cells through interaction with herpesvirus entry mediator. Nature Immunology. 2008;9(2):176–185. doi: 10.1038/ni1554. [DOI] [PubMed] [Google Scholar]
- 84.Wherry EJ, Blattman JN, Murali-Krishna K, Van Der Most R, Ahmed R. Viral persistence alters CD8 T-cell immunodominance and tissue distribution and results in distinct stages of functional impairment. Journal of Virology. 2003;77(8):4911–4927. doi: 10.1128/JVI.77.8.4911-4927.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sharpe AH, Wherry EJ, Ahmed R, Freeman GJ. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nature Immunology. 2007;8(3):239–245. doi: 10.1038/ni1443. [DOI] [PubMed] [Google Scholar]
- 86.Zajac AJ, Blattman JN, Murali-Krishna K, et al. Viral immune evasion due to persistence of activated T cells without effector function. Journal of Experimental Medicine. 1998;188(12):2205–2213. doi: 10.1084/jem.188.12.2205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Velu V, Titanji K, Zhu B, et al. Enhancing SIV-specific immunity in vivo by PD-1 blockade. Nature. 2009;458(7235):206–210. doi: 10.1038/nature07662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Day CL, Kaufmann DE, Kiepiela P, et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature. 2006;443(7109):350–354. doi: 10.1038/nature05115. [DOI] [PubMed] [Google Scholar]
- 89.Das A, Hoare M, Davies N, et al. Functional skewing of the global CD8 T cell population in chronic hepatitis B virus infection. Journal of Experimental Medicine. 2008;205(9):2111–2124. doi: 10.1084/jem.20072076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Urbani S, Amadei B, Tola D, et al. PD-1 expression in acute hepatitis C virus (HCV) infection is associated with HCV-specific CD8 exhaustion. Journal of Virology. 2006;80(22):11398–11403. doi: 10.1128/JVI.01177-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Nakamoto N, Cho H, Shaked A, et al. Synergistic reversal of intrahepatic HCV-specific CD8 T cell exhaustion by combined PD-1/CTLA-4 blockade. PLoS Pathogens. 2009;5(2) doi: 10.1371/journal.ppat.1000313. Article ID e1000313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kozako T, Yoshimitsu M, Fujiwara H, et al. PD-1/PD-L1 expression in human T-cell leukemia virus type 1 carriers and adult T-cell leukemia/lymphoma patients. Leukemia. 2009;23(2):375–382. doi: 10.1038/leu.2008.272. [DOI] [PubMed] [Google Scholar]
- 93.Lee PP, Yee C, Savage PA, et al. Characterization of circulating T cells specific for tumor-associated antigens in melanoma patients. Nature Medicine. 1999;5(6):677–685. doi: 10.1038/9525. [DOI] [PubMed] [Google Scholar]
- 94.Akbar AN, Henson SM. Are senescence and exhaustion intertwined or unrelated processes that compromise immunity? Nature Reviews Immunology. 2011;11(4):289–295. doi: 10.1038/nri2959. [DOI] [PubMed] [Google Scholar]
- 95.Ishida Y, Agata Y, Shibahara K, Honjo T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO Journal. 1992;11(11):3887–3895. doi: 10.1002/j.1460-2075.1992.tb05481.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Barber DL, Wherry EJ, Masopust D, et al. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439(7077):682–687. doi: 10.1038/nature04444. [DOI] [PubMed] [Google Scholar]
- 97.Petrovas C, Casazza JP, Brenchley JM, et al. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. Journal of Experimental Medicine. 2006;203(10):2281–2292. doi: 10.1084/jem.20061496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Harari A, Cellerai C, Enders FB, et al. Skewed association of polyfunctional antigen-specific CD8 T cell populations with HLA-B genotype. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(41):16233–16238. doi: 10.1073/pnas.0707570104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Trautmann L, Janbazian L, Chomont N, et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nature Medicine. 2006;12(10):1198–1202. doi: 10.1038/nm1482. [DOI] [PubMed] [Google Scholar]
- 100.Boni C, Fisicaro P, Valdatta C, et al. Characterization of hepatitis B virus (HBV)-specific T-cell dysfunction in chronic HBV infection. Journal of Virology. 2007;81(8):4215–4225. doi: 10.1128/JVI.02844-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Blank C, Kuball J, Voelkl S, et al. Blockade of PD-L1 (B7-H1) augments human tumor-specific T cell responses in vitro . International Journal of Cancer. 2006;119(2):317–327. doi: 10.1002/ijc.21775. [DOI] [PubMed] [Google Scholar]
- 102.Gehring AJ, Ho ZZ, Tan AT, et al. Profile of tumor antigen-specific cd8 T cells in patients with hepatitis B virus-related hepatocellular carcinoma. Gastroenterology. 2009;137(2):682–690. doi: 10.1053/j.gastro.2009.04.045. [DOI] [PubMed] [Google Scholar]
- 103.Ahmadzadeh M, Johnson LA, Heemskerk B, et al. Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood. 2009;114(8):1537–1544. doi: 10.1182/blood-2008-12-195792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Mumprecht S, Schürch C, Schwaller J, Solenthaler M, Ochsenbein AF. Programmed death 1 signaling on chronic myeloid leukemia-specific T cells results in T-cell exhaustion and disease progression. Blood. 2009;114(8):1528–1536. doi: 10.1182/blood-2008-09-179697. [DOI] [PubMed] [Google Scholar]
- 105.Zhang L, Gajewski TF, Kline J. PD-1/PD-L1 interactions inhibit antitumor immune responses in a murine acute myeloid leukemia model. Blood. 2009;114(8):1545–1552. doi: 10.1182/blood-2009-03-206672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Trautmann L, Janbazian L, Chomont N, et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nature Medicine. 2006;12(10):1198–1202. doi: 10.1038/nm1482. [DOI] [PubMed] [Google Scholar]
- 107.Blackburn SD, Shin H, Haining WN, et al. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nature Immunology. 2009;10(1):29–37. doi: 10.1038/ni.1679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Richter K, Agnellini P, Oxenius A. On the role of the inhibitory receptor LAG-3 in acute and chronic LCMV infection. International Immunology. 2009;22(1):13–23. doi: 10.1093/intimm/dxp107. [DOI] [PubMed] [Google Scholar]
- 109.Wherry EJ, Ha SJ, Kaech SM, et al. Molecular signature of CD8+ T cell exhaustion during chronic viral infection. Immunity. 2007;27(4):670–684. doi: 10.1016/j.immuni.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 110.Jones RB, Ndhlovu LC, Barbour JD, et al. Tim-3 expression defines a novel population of dysfunctional T cells with highly elevated frequencies in progressive HIV-1 infection. Journal of Experimental Medicine. 2008;205(12):2763–2779. doi: 10.1084/jem.20081398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Golden-Mason L, Palmer BE, Kassam N, et al. Negative immune regulator Tim-3 is overexpressed on T cells in hepatitis C virus infection and its blockade rescues dysfunctional CD4+ and CD8+ T cells. Journal of Virology. 2009;83(18):9122–9130. doi: 10.1128/JVI.00639-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.McMahan RH, Golden-Mason L, Nishimura MI, et al. Tim-3 expression on PD-1+ HCV-specific human CTLs is associated with viral persistence, and its blockade restores hepatocyte-directed in vitro cytotoxicity. Journal of Clinical Investigation. 2010;120(12):4546–4557. doi: 10.1172/JCI43127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Vali B, Jones RB, Sakhdari A, et al. HCV-specific T cells in HCV/HIV co-infection show elevated frequencies of dual Tim-3/PD-1 expression that correlate with liver disease progression. European Journal of Immunology. 2010;40(9):2493–2505. doi: 10.1002/eji.201040340. [DOI] [PubMed] [Google Scholar]
- 114.Fourcade J, Sun Z, Benallaoua M, et al. Upregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8+ T cell dysfunction in melanoma patients. Journal of Experimental Medicine. 2010;207(10):2175–2186. doi: 10.1084/jem.20100637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Sakuishi K, Apetoh L, Sullivan JM, Blazar BR, Kuchroo VK, Anderson AC. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. Journal of Experimental Medicine. 2010;207(10):2187–2194. doi: 10.1084/jem.20100643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Contardi E, Palmisano GL, Tazzari PL, et al. CTLA-4 is constitutively expressed on tumor cells and can trigger apoptosis upon ligand interaction. International Journal of Cancer. 2005;117(4):538–550. doi: 10.1002/ijc.21155. [DOI] [PubMed] [Google Scholar]
- 117.Boasso A, Vaccari M, Hryniewicz A, et al. Regulatory T-cell markers, indoleamine 2,3-dioxygenase, and virus levels in spleen and gut during progressive simian immunodeficiency virus infection. Journal of Virology. 2007;81(21):11593–11603. doi: 10.1128/JVI.00760-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Kaufmann DE, Kavanagh DG, Pereyra F, et al. Upregulation of CTLA-4 by HIV-specific CD4+ T cells correlates with disease progression and defines a reversible immune dysfunction. Nature Immunology. 2007;8(11):1246–1254. doi: 10.1038/ni1515. [DOI] [PubMed] [Google Scholar]
- 119.Kaufmann DE, Walker BD. PD-1 and CTLA-4 inhibitory cosignaling pathways in HIV infection and the potential for therapeutic intervention. Journal of Immunology. 2009;182(10):5891–5897. doi: 10.4049/jimmunol.0803771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Curran MA, Kim M, Montalvo W, Al-Shamkhani A, Allison JP. Combination CTLA-4 blockade and 4-1BB activation enhances tumor rejection by increasing T-cell infiltration, proliferation, and cytokine production. PLoS ONE. 2011;6(4) doi: 10.1371/journal.pone.0019499. Article ID e19499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Derre L, Rivals JP, Jandus C, et al. BTLA mediates inhibition of human tumor-specific CD8+ T cells that can be partially reversed by vaccination. Journal of Clinical Investigation. 2010;120(1):157–167. doi: 10.1172/JCI40070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Xu XS, Zhang Z, Gu LL, Wang FS. [BTLA Characterization and its association with disease progression in patients with chronic HIV-1 infection.] Chinese Journal of Cellular and Molecular Immunology. 2009;25:1158–1160. [PubMed] [Google Scholar]
- 123.Zhang Z, Xu X, Lv J, et al. B and T lymphocyte attenuator down-regulation by HIV-1 depends on type I interferon and contributes to T-cell hyperactivation. Journal of Infectious Diseases. 2011;203(11):1668–1678. doi: 10.1093/infdis/jir165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Antonelli LRV, Mahnke Y, Hodge JN, et al. Elevated frequencies of highly activated CD4+ T cells in HIV+ patients developing immune reconstitution inflammatory syndrome. Blood. 2010;116(19):3818–3827. doi: 10.1182/blood-2010-05-285080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Matsuzaki J, Gnjatic S, Mhawech-Fauceglia P, et al. Tumor-infiltrating NY-ESO-1-specific CD8+ T cells are negatively regulated by LAG-3 and PD-1 in human ovarian cancer. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(17):7875–7880. doi: 10.1073/pnas.1003345107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Peritt D, Sesok-Pizzini DA, Schretzenmair R, et al. C1.7 antigen expression on CD8+ T cells is activation dependent: increased proportion of C1.7+CD8+ T cells in HIV-1-infected patients with progressing disease. Journal of Immunology. 1999;162(12):7563–7568. [PubMed] [Google Scholar]
- 127.Aldy KN, Horton NC, Mathew PA, Mathew SO. 2B4+ CD8+ T cells play an inhibitory role against constrained HIV epitopes. Biochemical and Biophysical Research Communications. 2011;405(3):503–507. doi: 10.1016/j.bbrc.2011.01.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Williams H, Macsween K, McAulay K, et al. Analysis of immune activation and clinical events in acute infectious mononucleosis. Journal of Infectious Diseases. 2004;190(1):63–71. doi: 10.1086/421276. [DOI] [PubMed] [Google Scholar]
- 129.Bengsch B, Seigel B, Ruhl M, et al. Coexpression of PD-1, 2B4, CD160 and KLRG1 on exhausted HCV-specific CD8+T cells is linked to antigen recognition and T cell differentiation. PLoS Pathogens. 2010;6(6) doi: 10.1371/journal.ppat.1000947. Article ID e1000947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Raziorrouh B, Ulsenheimer A, Schraut W, et al. Inhibitory molecules that regulate expansion and restoration of HCV-specific CD4+ T cells in patients with chronic infection. Gastroenterology. 2011;141(4):1422–1431. doi: 10.1053/j.gastro.2011.07.004. [DOI] [PubMed] [Google Scholar]
- 131.Yamamoto T, Price DA, Casazza JP, et al. Surface expression patterns of negative regulatory molecules identify determinants of virus-specific CD8+ T-cell exhaustion in HIV infection. Blood. 2011;117(18):4805–4815. doi: 10.1182/blood-2010-11-317297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Kassu A, Marcus RA, D’Souza MB, et al. Regulation of virus-specific CD4+ T cell function by multiple costimulatory receptors during chronic HIV infection. Journal of Immunology. 2010;185(5):3007–3018. doi: 10.4049/jimmunol.1000156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Lipson EJ, Drake CG. Ipilimumab: an anti-CTLA-4 antibody for metastatic melanoma. Clinical Cancer Research. 2011;17(22):6958–6962. doi: 10.1158/1078-0432.CCR-11-1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Danielli R, Ridolfi R, Chiarion-Sileni V, et al. Ipilimumab in pretreated patients with metastatic uveal melanoma: safety and clinical efficacy. Cancer Immunology, Immunotherapy. 2012;61(1):41–48. doi: 10.1007/s00262-011-1089-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Brahmer JR, Drake CG, Wollner I, et al. Phase I study of single-agent anti-programmed death-1 (MDX-1106) in refractory solid tumors: safety, clinical activity, pharmacodynamics, and immunologic correlates. Journal of Clinical Oncology. 2010;28(19):3167–3175. doi: 10.1200/JCO.2009.26.7609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Kline J, Gajewski TF. Clinical development of mAbs to block the PD1 pathway as an immunotherapy for cancer. Current Opinion in Investigational Drugs. 2010;11(12):1354–1359. [PubMed] [Google Scholar]
- 137.Hodi FS, Mihm MC, Soiffer RJ, et al. Biologic activity of cytotoxic T lymphocyte-associated antigen 4 antibody blockade in previously vaccinated metastatic melanoma and ovarian carcinoma patients. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(8):4712–4717. doi: 10.1073/pnas.0830997100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Phan GQ, Yang JC, Sherry RM, et al. Cancer regression and autoimmunity induced by cytotoxic T lymphocyte-associated antigen 4 blockade in patients with metastatic melanoma. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(14):8372–8377. doi: 10.1073/pnas.1533209100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Hryniewicz A, Boasso A, Edghill-Smith Y, et al. CTLA-4 blockade decreases TGF-β, IDO, and viral RNA expression in tissues of SIVmac251-infected macaques. Blood. 2006;108(12):3834–3842. doi: 10.1182/blood-2006-04-010637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Cecchinato V, Tryniszewska E, Ma ZM, et al. Immune activation driven by CTLA-4 blockade augments viral replication at mucosal sites in simian immunodeficiency virus infection. Journal of Immunology. 2008;180(8):5439–5447. doi: 10.4049/jimmunol.180.8.5439. [DOI] [PMC free article] [PubMed] [Google Scholar]